Fecundação
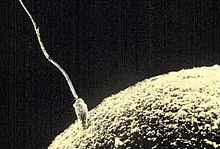
Momento que o espermatozoide começa a penetrar no ovócito II
Em Biologia, chama-se fecundação, conceção ou fertilização ao processo em que um espermatozoide penetra no óvulo, como no caso dos seres humanos e da maioria dos mamíferos, em outras espécies (nos restantes animais)[1], ou em que o tubo polínico penetra no óvulo das plantas durante o processo de reprodução[2]. Confrontar com singamia e conjugação.
Índice
1 Fecundação entre as plantas
2 Fecundação entre animais
3 Reconhecimento do óvulo e espermatozoide
4 Herança matrilinear mitocondrial
5 Fecundação em ouriços do mar
6 Fecundação em mamíferos
7 Fecundação na espécie humana
8 Fecundação em Anfíbios (modelo: Xenopus)
9 Referências
Fecundação entre as plantas |
No caso das plantas com semente, deve-se diferenciar o fenômeno da fecundação propriamente dito (união íntima de duas células sexuais até confundirem seus respectivos núcleos e, em maior ou menor grau, seus citoplasmas),[3] do processo biológico que o antecede - a polinização, no qual os grãos de pólen, desenvolvidos nas tecas que contém cada antera de um estame (órgão reprodutora masculino), são transportados pelo vento ou por insetos até os estigmas, onde germinan emitindo um tubo polínico que cresce até o ovario. Nesse caso não se trata de gametas, mas de esporos, pois cada grão de pólen contém dois gametas ou células reprodutoras masculinas, que são transportadas a um órgão reprodutor feminino (carpelo) de outra flor (polinização cruzada) ou da misma flor (Autopolinización).
Os espermatozoides, depois do ato sexual, movimentam-se em direção às tubas uterinas. Os líquidos nutritivos do esperma e um muco do sistema reprodutor feminino facilitam o seu movimento em direção ao ovócito II, que foi libertado pelo ovário e se encontra numa das tubas uterinas.
Fecundação entre animais |
Em geral, a fecundação é intraespecífica, isto é, processa-se apenas entre indivíduos da mesma espécie. Mas há casos de fecundação interespecífica, entre indivíduos de duas espécies evolutivamente próximas. Os indivíduos que nascem destas fecundações designam-se por híbridos (exemplos: os cavalos (Eqqus ferus caballus) e os burros (Eqqus africanus asinus) podem cruzar-se, dando a origem a um híbrido: o macho (burro, em português do Brasil) ou mula, que neste caso são estéreis; o jaguar e o leopardo podem cruzar, dando origem a um híbrido, que neste caso é fértil, etc.).
A célula reprodutora feminina (óvulo numas espécies ovócito II noutras) possui barreiras para a penetração dos espermatozoides: a corona radiata (mais externa, composta de células foliculares) e a zona pelúcida (camada glicoproteica situada após a corona radiata). Os espermatozoides, gametas masculinos, possuem na cabeça o acrossomo, que começa a liberar enzimas hidrolíticas ao entrar em contato com tais barreiras. Após vencê-las, ocorre a fusão entre as membranas dos dois gametas.
Imediatamente após a fecundação, as células foliculares glandulares que envolvem a célula reprodutora feminina retraem-se, liberta-se o conteúdo dos grânulos corticais formando a membrana de fecundação que não vai permitir a entrada de mais espermatozoides.
A fecundação alcança dois propósitos diferentes: sexo (a combinação de genes derivados de dois genitores) e reprodução (a geração de um novo organismo). Ela pode ser externa (gametas são liberados no ambiente) ou interna (ocorre no interior do organismo).
Embora os detalhes da fecundação variem em diferentes organismos, a concepção consiste geralmente de quatro eventos principais, que não estão relacionados apenas à fusão dos gametas:
1. Contato e reconhecimento entre óvulo e espermatozoide, geralmente garantindo que ambos sejam da mesma espécie;
2. Regulação da entrada do espermatozoide no óvulo, garantindo que apenas um pronúcleo masculino possa se fundir com um pronúcleo feminino, evitando a poliespermia;
3. Fusão do material genético do espermatozoide e do óvulo;
4. Ativação do metabolismo do óvulo e início do desenvolvimento. [4]
Reconhecimento do óvulo e espermatozoide |
A interação dos gametas geralmente envolve cinco passos:
1. Quimioatração do espermatozoide por moléculas solúveis secretadas pelo óvulo;
2. Exocitose da vesícula acrossômica do espermatozoide e liberação de suas enzimas;
3. Ligação do espermatozoide à matriz extracelular do óvulo (envelope vitelínico ou zona pelúcida);
4. Passagem do espermatozoide pela matriz extracelular;
5. Fusão das membranas dos gametas. [4]
Herança matrilinear mitocondrial |
Todas as mitocôndrias, organelas responsáveis pela respiração celular, são herdadas da mãe do indivíduo. Isto ocorre porque as mitocôndrias do espermatozoide se encontram na parte da cauda. Ao fecundar o ovócito II, o espermatozoide emparelha de lado (equatorialmente) a membrana do ovócito II e então digere a membrana do Ovócito II, onde substancias presentes dentro do Ovócito II irão ajudar a "sugar" somente o pró-núcleo do espermatozoide para dentro que contém a informação genética, era o que dizia algumas literaturas antigas. Porém foi descoberto que o espermatozoide entra todo para dentro do ovócito, mas que depois a cauda é degenerada no interior do ovócito II.
As mitocôndrias presentes na cauda, como a cauda é degenerada,não são herdadas pelo embrião. Estas mitocôndrias servem apenas para ajudar no movimento do espermatozoide até ao ovócito II. A partir das mitocôndrias, também é possível realizar um teste de "DNA mitocondrial", para provar a maternidade.
Fecundação em ouriços do mar |
Ouriços do mar têm fecundação externa, liberando seus gametas na água. Esse ambiente é muito diluído e compartilhado por outras espécies que também liberam seus gametas. Para garantir que seus gametas se encontrem, e que não fertilizem ou sejam fertilizados por gametas de outras espécies, além de liberar uma grande quantidade de gametas existem também outros mecanismos.
Um deles é a atração espécie-específica dos espermatozoides, que já foi documentada também em várias outras espécies, como cnidários, moluscos, equinodermos, anfíbios e urocordados. Em muitas espécies, ocorre a atração dos espermatozoides por quimiotaxia (seguindo um gradiente químico secretado pelo óvulo). Há casos, como o do cnidário Orthopyxis caliculata, em que o óvulo, além de secretar um fator quimiotático, também controle o momento da liberação, garantindo que este fator só seja liberado quando o gameta feminino está pronto para ser fecundado. Os mecanismos de quimiotaxia diferem entre espécies, assim como diferem as moléculas quimiotáticas, mesmo em espécies próximas, garantindo a especificidade para cada espécie.
Em ouriços do mar, os espermatozoides adquirem motilidade ao serem lançados na água do mar. Nos testículos, eles não são capazes de se mover devido ao menor pH (cerca de 7,2) da gônada, que é mantido por altas concentrações de CO2. Ao entrar em contato com a água do mar, o pH do espermatozoide sobe para 7,6, ativando a ATPase da dineína, o que permite a movimentação do flagelo. No entanto, para que esse movimento tenha direção, em equinodermos, são necessários peptídeos quimiotáticos chamados de peptídeos ativadores de espermatozoide (do inglês sperm-activating peptides, ou SAPs). Um exemplo de SAP é resact um peptídeo de 14 aminoácidos, específico de A. punctulata. Espermatozoides desta espécie tem receptores de membrana que se ligam a resact, e essa ligação por sua vez ativa a enzima guanilil ciclase no lado citoplasmático do receptor, levando à maior produção de GMP cíclico (cGMP), que ativa canais de cálcio na membrana plasmática da cauda, levando ao influxo destes íons (Ca2+) para dentro da cauda, permitindo a percepção pelo espermatozoide do gradiente de resact. Resact também age como ativador do espermatozoide. O aumento de cGMP e Ca2+ também ativa o aparato gerador de ATP nas mitocôndrias, dando energia para a movimentação.
Uma segunda interação entre a camada gelatinosa do óvulo e o espermatozoide é a reação acrossômica. Na maioria dos invertebrados marinhos, a reação acrossômica tem dois componentes: a fusão do acrossomo com a membrana do espermatozoide (uma exocitose), que libera enzimas e proteassomos (complexos que digiram proteínas) do acrossomo, digerindo assim a camada gelatinosa do óvulo, e a extensão do processo acrossômico, que ao atingir a superfície do óvulo adere ao envelope vitelínico ligando óvulo e espermatozoide. Em ouriços do mar, essa reação é iniciada com a ligação de polissacarídeos contendo sulfato da camada gelatinosa do óvulo a receptores acima da vesícula acrossomal na membrana do espermatozoide, sendo que esses polissacarídeos também são espécie-específicos.
Por fim, após a reação acrossomal, outro evento ocorre: a ligação do espermatozoide à superfície do óvulo. Em ouriços do mar, isso ocorre por meio de uma proteína chamada bindin, que também varia de forma espécie-específica. Acredita-se que bindin se ligue a receptores do envelope vitelínico, que ficam agregados em complexos, o que explicaria porque a ligação de espermatozoides não ocorre em toda a superfície do óvulo mesmo em condições de saturação.
Bindin e outras proteínas de reconhecimento entre gametas estão entre as proteínas conhecidas que evoluem mais rapidamente, e acredita-se que a coevolução entre proteínas de reconhecimento de gametas femininos e masculinos é importante para a geração de barreiras reprodutivas que levem à especiação.
Após o reconhecimento, ocorre a fusão dos gametas. Para isso, a actina presente no óvulo se polimeriza, formando o cone de fertilização (processo homólogo ao processo acrossômico), criando uma ponte citoplasmática pela qual o núcleo e a cauda do espermatozoide passam e entram no óvulo. Essa fusão é um processo ativo, mediada por proteínas “fusogênicas”, que em ouriços do mar, é um papel também desempenhado por bindin.
Após a fusão do primeiro espermatozoide, a fusibilidade da membrana do óvulo passa a ser um problema, pois poderia levar à entrada de mais espermatozoides levando à poliespermia, que leva à morte ou ao desenvolvimento anormal. Para evitar que isso ocorra, diferentes mecanismos evoluíram, dois deles ocorrendo em ouriços do mar. Um é o bloqueio rápido à poliespermia, em que a membrana tem seu potencial de repouso (de -70 mV) alterado para um potencial positivo (de +20 mV), devido à abertura de canais de sódio e o influxo de Na+ no óvulo. Não se sabe se a abertura destes canais ocorre devido à ligação com o primeiro espermatozoide, ou devido à sua fusão com o óvulo. Também não se sabe porque a mudança de potencial leva ao bloqueio da entrada de outros espermatozoides, mas acredita-se que isso possa ocorrer devido a algum componente sensível a voltagem carregado pelo espermatozoide (por exemplo, uma proteína fusogênica carregada positivamente). Esse mecanismo, no entanto, é passageiro.
Assim, em seguida, ocorre o bloqueio lento à poliespermia, ou reação dos grânulos corticais, que remove os espermatozoides restantes ligados ao envelope vitelínico. Este é um mecanismo mais lento, ativo cerca de um minuto após a fusão, e é encontrado em ouriço do mar e na maioria dos mamíferos. Após a fusão dos gametas, íons Ca2+ são liberados do retículo endoplasmático do óvulo, e essa reação se propaga, pois o cálcio leva à liberação de mais cálcio. O cálcio liberado leva à fusão de grânulos corticais próximos à membrana com a membrana do óvulo e a liberação de seu conteúdo entre a membrana celular e as proteínas do envelope vitelínico. Várias proteínas são liberadas com essa reação. Uma delas é a protease serina grânulo cortical, que cliva a ligação entre as proteínas do envelope vitelínico e a membrana, e também retira os receptores de bindin e os espermatozoides ligados a eles. Os componentes dos grânulos corticais formam o envelope de fertilização, que começa a se formar no local de fusão e se propaga pelo resto da célula. Este envelope é elevado da membrana celular devido a mucopolissacarídeos, também liberados pelos grânulos corticais, que absorvem água e expandem o espaço entre a membrana e o envelope de fertilização. O envelope também é estabilizado por proteínas dos grânulos, e um quarto grupo de proteínas granulares corticais, incluindo hialina, formam uma camada sobre o óvulo, que estende suas microvilosidades, ligando-se à camada de hialina, que suportará os blastômeros durante a clivagem.
A liberação de cálcio do retículo endoplasmático também é responsável pela ativação do metabolismo e o início do desenvolvimento, liberando inibidores de mensagens estocadas no óvulo, permitindo que esses RNAs mensageiros sejam traduzidos, e também liberando a inibição da divisão nuclear, possibilitando a clivagem. A liberação de cálcio do retículo endoplasmático ocorre devido à via do inositol trifosfato (IP3). O fosfolipídio de membrana fosfatidilinosol bifosfato (PIP2) é clivado pela fosfolipase C (PLC) em dois compostos ativos: IP3 e diacilglicerol (DAG). IP3 libera Ca2+ no citoplasma ao abrir canais de cálcio no retículo endoplasmático, e DAG ativa a proteína quinase C, que ativa uma proteína que troca íons de sódio por íons de hidrogênio, aumentando o pH (essa troca também necessita de cálcio). Acredita-se que, em ouriços do mar, a ligação do espermatozoide ou a fusão ativa PLC por proteínas G e quinases da família Src. É também possível que componentes do espermatozoide, como o ácido nicotínico adenina dinucleotídeo fosfato (NAADP), também sirva como liberador de Ca2+ do retículo.
O influxo de Ca2+, além das reações já citadas também leva a outras respostas: dentre as primeiras, estão a ativação de reações metabólicas que iniciam o desenvolvimento, como a ativação da enzima NAD+quinase, que converte NDA+ a NADP+, que é essencial para a síntese de lipídios, o que é importante para a formação de novas membranas durante a clivagem. NADP+ também é usado para fazer NAADP. A liberação de cálcio também afeta o consumo de oxigênio, levando a um aumento na sua redução. Dentre reações mais tardias, estão a retomada da síntese de DNA, devido ao aumento do pH e ao Ca2+, que inativa a enzima MAP quinase, responsável pela inibição da síntese de DNA; e também a retomada da síntese de proteínas, devido à degradação de inibidores (como a proteína de ligação eIF4E) da tradução dos RNAs mensageiros estocados no óvulo.
Após a fusão de espermatozoide e óvulo, as mitocôndrias e o flagelo do espermatozoide são degradados. No entanto, o centrossomo necessário para a produção do fuso mitótico nas divisões subsequentes é derivado do centríolo do espermatozoide. A fecundação em ouriços do mar ocorre após a segunda divisão meiótica, ou seja, já há um pró-núcleo feminino. O núcleo do espermatozoide descondensa formando o pró-núcleo masculino. Para isso, o envelope nuclear desmancha, expondo a cromatina, que tem suas proteínas trocadas por proteínas do óvulo, permitindo a descondensação e a adesão ao envelope nuclear. [4]
Fecundação em mamíferos |
A fecundação de mamíferos é interna. A população de espermatozoides ejaculados é, em geral, heterogênea. A fecundação ocorre numa região do oviduto chamada ampola. O trato reprodutivo feminino regula ativamente o transporte e a maturação de ambos os gametas.
Um dos mecanismos para o transporte dos gametas até a ampola é a translocação. No caso do oócito, ele é liberado do ovário rodeado por uma matriz contendo células cumulus, que são necessárias para que as fímbrias do oviduto capturem o oócito. Em seguida, o oócito é transportado por meio de batimentos ciliares. Os espermatozoides, por sua vez, também sofrem translocação, sendo transportados para o oviduto por meio de contrações musculares uterinas.
Espermatozoides recém ejaculados não são capazes de sofrer reação acrossômica, ou fertilizar o oócito. Para que se tornem capazes, é preciso que residam um tempo no trato reprodutivo feminino, onde sofrem capacitação. Caso não sejam capacitados, eles não conseguem passar a matriz de células cumulus e alcançar o oócito. Cinco mudanças moleculares são consideradas importantes: a remoção de colesterol da membrana por proteínas albumina do trato reprodutivo feminino, que acredita-se levar a uma reorganização de rafts de lipídios, levando a proteínas importantes na ligação à zona pelúcida e na reação acrossômica a se localizarem na parte anterior da cabeça; remoção de proteínas ou carboidratos da superfície do espermatozoide, o que possivelmente leva a liberação de locais de ligação à zona pelúcida; o potencial de membrana se torna mais negativo devido à saída de íons de potássio, mudança que pode abrir canais de cálcio, íons necessários para a produção de AMP cíclico (cAMP) e para eventos de fusão de membrana e reação acrossômica; fosforilação de proteínas, em particular duas chaperonas que migram para a superfície da cabeça do espermatozoide quando fosforiladas, uma delas chamada Izumo, que é crítica para a fusão de espermatozoide e óvulo; a membrana acrossomal muda, se preparando para fusão.
Parece haver conexão entre translocação e capacitação. Foi documentado que espermatozoides incapacitados se ligam ativamente às membranas de células do oviduto que precedem a ampola (o istmo). Essa ligação parece ser quebrada com a capacitação, e assim, os espermatozoides são liberados aos poucos.
Diferentes regiões do trato reprodutivo secretam diferentes moléculas que influenciam a capacitação e também a motilidade. Durante a capacitação, os espermatozoides se tornam hiperativados, nadando a maior velocidade e com mais força, devido à abertura de canais de cálcio na cauda, liberando o espermatozoide de sua ligação ao oviduto, e permitindo (juntamente com uma enzima hialuronidase) que o espermatozoide digira um caminho pela matriz de células cumulus. Ao chegar próximo à ampola, espermatozoides capacitados são direcionados à ampola pelo gradiente de calor (que é maior na ampola), e pelo gradiente quimiotático, ao chegar mais próximos ao oócito. Uma das moléculas responsáveis pela quimiotaxia é a progesterona, que ativa canais de cálcio na cauda, levando à hiperativação.
Antes de se ligar ao oócito, o espermatozoide deve se ligar à zona pelúcida, que tem um papel análogo ao envelope vitelínico de invertebrados. A zona pelúcida é composta de três glicoproteínas principais ZP1, ZP2 e ZP3 (do inglês, zone proteins), além de outras proteínas acessórias. A ligação à zona pelúcida é relativamente, mas não absolutamente, espécie-específica. Hoje, acredita que espermatozoides bem-sucedidos sofrem a reação acrossômica ao atingir às células cumulus, antes de chegar à zona pelúcida.
Em mamíferos, o contato com o óvulo ocorre com a parte lateral da cabeça do espermatozoide, e a fusão ocorre em um local da membrana que fazia parte do interior do acrossomo, a região equatorial. Uma proteína do oócito que participa da fusão é CD9, e uma proteína do espermatozoide é Izumo. Não se sabe se elas se ligam uma a outra, e há ainda outras proteínas que são candidatas à participação na fusão dos gametas. Após a fusão, ao contrário de ouriços do mar, não ocorre o bloqueio rápido contra poliespermia. No entanto, ocorre o bloqueio lento, com a fusão de grânulos corticais e clivagem de proteínas de ZP2.
O pró-núcleo masculino tem seu DNA enrolado por protaminas, que está compactada por pontes dissulfeto, que são reduzidas por glutationa do oócito, permitindo a descondensação. A fecundação em mamíferos ocorre com o pró-núcleo feminino ainda em metáfase II. As oscilações de cálcio resultantes da entrada do espermatozoide levam à ativação de uma quinase que leva à proteólise de ciclina e securina, retomando a meiose. A síntese de DNA ocorre separadamente nos dois pró-núcleos. O centrossomo é derivado do espermatozoide. Os microtúbulos unem os dois pró-núcleos, cujos envelopes nucleares se quebram ao se encontrarem, e os cromossomos já se condensam e se organizam no fuso mitótico. Assim, um núcleo diploide verdadeiro só é visto no estágio de duas células. As mitocôndrias paternas são destinadas à destruição.
A fecundação em mamíferos também leva à liberação de cálcio, pela produção de IP3 a partir de PLC. No entanto, no caso de mamíferos, parece que a PLC está não na membrana do oócito, mas na cabeça do espermatozoide. O cálcio tem o efeito de ativação do oócito, levando ao início do metabolismo e desenvolvimento. Parece também haver necessidade de Ca2+ extracelular. [4]
Fecundação na espécie humana |
Se a mulher estiver fora do período fértil os espermatozoides geralmente não têm qualquer possibilidade de entrar sequer no útero, visto existir um muco cervical muito rico em fibras que impossibilita a entrada de espermatozoides. Mas se ela se encontrar no período fértil o colo do útero para além de se encontrar mais aberto vai conter um muco cervical mais fluido e com a rede de fibras mais aberta permitindo que os espermatozoides entrem (cerca de 1% dos espermatozoides contidos no ejaculado) estes dirigem-se para as trompas ao encontro do ovócito II, ocorrendo mais tarde a fusão de um deles com o ovócito II após ter conseguido atravessar a zona pelúcida. Quando isto acontece, as células foliculares glandulares que envolvem o ovócito II retraem-se e o ovócito II completa a divisão II da meiose. Ao mesmo tempo liberta-se o conteúdo dos grânulos corticais formando a membrana de fecundação que não vai permitir a entrada de mais espermatozoides.
Após estar completa a divisão II da meiose e formado o óvulo os núcleos dos dois gametas fundem-se (cariogamia) e forma-se o ovo, com uma totalmente nova associação de genes e que vai caracterizar o indivíduo por toda a sua vida do ponto de vista genético.
Seguidamente e ocorre a fase embrionária (com uma duração aproximada de 16 semanas) que se inicia ainda na trompa com a divisão celular (por mitose) dando origem à formação do embrião, ao mesmo tempo em que se dirige para o útero. Para que este mantenha o endométrio em estado de desenvolvimento conveniente para o embrião, tem que continuar a receber hormônios ovários, mas como a LH vai acabar por deixar de ser produzida devido ao retrocontrolo negativo, o próprio embrião produz o hormônio gonadotrópico (HCG) que vai impedir a regressão do corpo amarelo e manter a produção de progesterona e estrogênios.
A manutenção da produção destes hormônios continua a inibir o hipotálamo de produzir GnRH e por consequência manter interrompido o ciclo éstrico (ciclo ovárico e ciclo uterino). Chegado ao útero, a zona pelúcida que envolve o embrião é destruída e este começa a crescer em virtude do fornecimento de nutrientes pelas glândulas do Endométrio. O embrião vai começar a afundar no endométrio por ação de enzimas que liberta sendo ao mesmo tempo envolvido por outras células deste — nidação. Após este fenômeno começam a formar-se as estruturas embrionárias (placenta, cordão umbilical, saco amniótico que vai conter o líquido amniótico que serve de proteção ao novo ser).
Por volta da quinta semana a placenta passa a produzir ela própria os estrogênios e a progesterona para manter o endométrio, deixando de ser produzido a HCG o que leva à regressão do corpo amarelo. Ocorre igualmente a produção de um hormônio pela placenta que leva à preparação das glândulas mamárias para o aleitamento. Esta fase termina quando estiverem esboçados os diferentes órgãos do novo indivíduo.
Segue-se a fase fetal em que o que vai ocorrer essencialmente é o crescimento e maturação dos órgãos o que termina aproximadamente ao fim de 40 semanas, seguindo-se o nascimento.
Fecundação em Anfíbios (modelo: Xenopus) |
O ovo de Xenopus é circular e possui um polo animal e um polo vegetal, citoplasma cortical (pigmentado no polo animal e não pigmentado no polo vegetal) e citoplasma subcortical. A fecundação é externa e a entrada do espermatozoide no ovo ocorre sempre no polo animal, sendo a região de entrada do espermatozoide definida como ventral. Assim que a fecundação ocorre, o citoplasma cortical gira 30 graus na direção de entrada do espermatozoide, enquanto o citoplasma subcortical permanece imóvel, e portanto, com uma pequena região agora exposta e denominada grey crescent.
A rotação cortical do citoplasma do ovo de Xenopus possui algumas funções biológicas, sendo elas: reorganização dos microtúbulos abaixo do citoplasma cortical, especificação do eixo dorso-central e exposição do citoplasma subcortical (área definida como grey crescent).
O ovo também possui proteínas e RNAs mensageiros herdados da mãe que são chamados determinantes maternos. Estes determinantes são assim chamados por serem herdados da mãe e por agirem no começo do desenvolvimento do embrião, enquanto este não tem seu genoma completamente ativado.
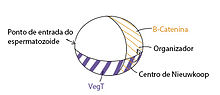
Alguns destes determinantes maternos são encontrados através de todo o ovo em forma de proteínas, como GSK3 e β-catenina. Outros determinantes maternos são encontrados apenas no polo vegetal em forma de proteínas (Dsh, GBP, Wnt11) ou RNAs mensageiros (VegT, Vg1). Estes determinantes farão parte da via de sinalização Wnt ou TGFβ, sendo a primeira responsável pelo estabelecimento da região dorsal do organismo e a segunda pela especificação da mesoderme.
Blástula de xenopus
Via de sinalização Wnt: os determinantes maternos Dsh, GBP e Wnt11 estão localizados no citoplasma cortical do polo vegetal. Ao ocorrer a rotação cortical após a fecundação, estes determinantes também mudam de lugar, sendo encontrados agora 30 graus no sentido anti-horário do polo vegetal em relação à posição anterior. A expressão destes determinantes inibirá a ação de GSK3 na região dorsal do ovo. GSK3 inibe a ação da β-catenina, no entanto, como ele está inibido, a β-catenina está livre e consegue se deslocar para o núcleo das células da região dorsal do ovo.
A presença de β-catenina na região dorsal do ovo induz a formação do Centro de Nieuwkoop, também chamado de organizador. O Centro de Nieuwkoop irá secretar a proteína Nodal que irá definir as células desta região como mesoderme dorsal. Ou seja, a rotação cortical especifica o eixo dorso-ventral ao tornar β-catenina disponível para agir como um fator de transcrição no núcleo, levando à ativação do Centro de Nieuwkoop que secreta a proteína Nodal e estabelece a identidade da mesoderme dorsal.
Via de sinalização TGFβ: na região ventral por sua vez, não há expressão de Dsh, GBP ou Wnt11, logo, GSK3 está livre e consegue inibir a β-catenina, levando à sua degradação. Vg1 e VegT são expressos na região ventral, induzindo baixos níveis da proteína Nodal, o que ela à especificação das células dessa região como mesoderme ventral.[5][6][7]
Referências
↑ Bruna, Iara, Michele, Vanessa e Dinailde - 1º O “Gametogênese animal” no site do Instituto de Educação Socioambiental do Brasil acessado a 10 de agosto de 2009
↑ “Os Seres Vivos - Reino das Plantas -O Angiospermas - A Flor” no site PortalBrasil.net acessado a 10 de agosto de 2009
↑ Font Quer, P. 1977. Diccionario de botánica. Barcelona: Ed. Labor. p. 459.
↑ abcd GILBERT., Scott F. (2014). Developmental Biology. [S.l.]: Swathmore College and the University of Helsinki
↑ Larabell, C. A., Torres, M., Rowning, B. A., Yost, C., Miller, J. R., Wu, M., ... & Moon, R. T. (1997). Establishment of the dorso-ventral axis in Xenopus embryos is presaged by early asymmetries in β-catenin that are modulated by the Wnt signaling pathway. The Journal of cell biology, 136(5), 1123-1136.
↑ Miller, J. R., Rowning, B. A., Larabell, C. A., Yang-Snyder, J. A., Bates, R. L., & Moon, R. T. (1999). Establishment of the Dorsal–Ventral Axis inXenopus Embryos Coincides with the Dorsal Enrichment of Dishevelled That Is Dependent on Cortical Rotation. The Journal of cell biology, 146(2), 427-438.
↑ Tickle, C. (2011). Principles of development. Oxford university press.


